Graduate School of Agriculture
Kindai University
Kindai University
尋媶撪梕Research
怉暔偺柶塽婡峔偺奣梫
 丂怉暔偼丄昦尨嬠擣幆庴梕懱乮昦尨嬠擣幆僙儞僒乕乯
傪傕偪丄昦尨嬠偺姶愼傪専弌偟偰恦懍側杊屼墳摎傪桿摫偟傑偡丅偙偺傛偆側庴梕懱傪夘偟偨杊屼斀墳偺桿摫婡峔偼丄
摦暔偺愭揤惈帺慠柶塽偲旕忢偵傛偔帡偰偄傞偨傔丄嬤擭丄乽怉暔柶塽 (Plant Immunity)乿偲屇偽傟偰偄傑偡丅
丂怉暔偼丄昦尨嬠擣幆庴梕懱乮昦尨嬠擣幆僙儞僒乕乯
傪傕偪丄昦尨嬠偺姶愼傪専弌偟偰恦懍側杊屼墳摎傪桿摫偟傑偡丅偙偺傛偆側庴梕懱傪夘偟偨杊屼斀墳偺桿摫婡峔偼丄
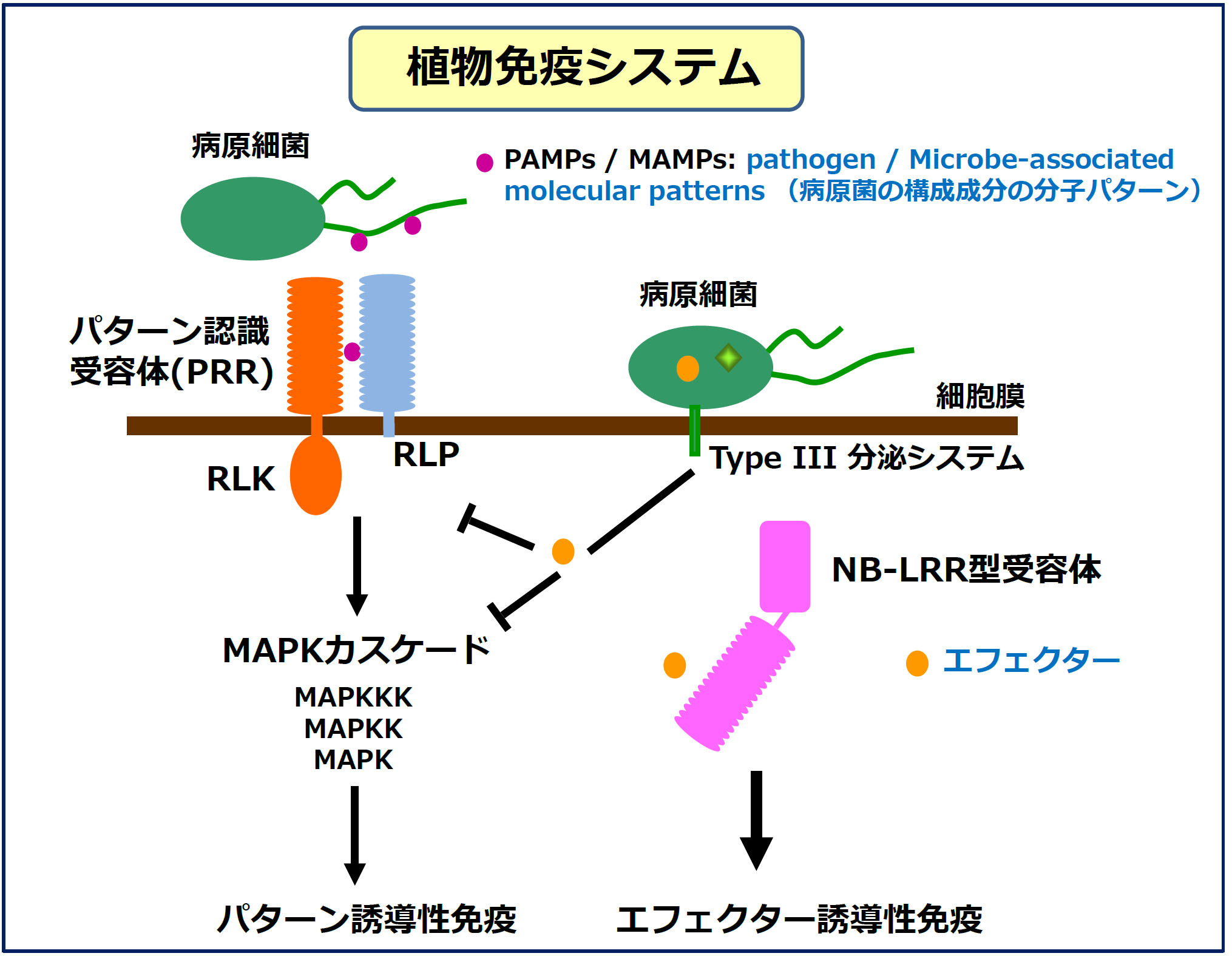
摦暔偺愭揤惈帺慠柶塽偲旕忢偵傛偔帡偰偄傞偨傔丄嬤擭丄乽怉暔柶塽 (Plant Immunity)乿偲屇偽傟偰偄傑偡丅丂昦尨嬠擣幆庴梕懱偼丄峔憿揑摿挜偐傜戝偒偔2庬椶丄嵶朎枌嬊嵼宆偲嵶朎撪偵懚嵼偡傞NB-LRR宆乮徻嵶偼屻弎乯偵暘偗傜傟傑偡(恾侾)丅 嵶朎枌偵懚嵼偡傞庴梕懱偼丄僷僞乕儞擣幆庴梕懱乮Pattern Recognition Receptor (PRR)乯偲屇偽傟丄嵶朎奜偵昦尨嬠/旝惗暔偺峔惉惉暘(PAMP/MAMP: Pathogen/Microbe-Associated Molecular Pattern)傪専弌偡傞僪儊僀儞傪帩偪傑偡丅PRR偺嵶朎奜僪儊僀儞偲偟偰偼丄LRR (leucine-rich repeat)傗LysM (lysine rich motif)側偳偑抦傜傟偰偄傑偡丅偝傜偵丄PRR偺嵶朎撪僪儊僀儞偵僾儘僥傿儞僉僫乕僛僪儊僀儞傪帩偮傕偺偼丄Receptor-like kinase(RLK)偲屇偽傟 丄帩偨側偄傕偺偼丄Receptor-like protein (RLP)偲屇偽傟偰偄傑偡丅PRR偑PAMPs/MAMPs傪専弌偟偰桿摫偡傞柶塽墳摎偼丄僷僞乕儞桿摫惈柶塽 (Pattern-triggered immunity乯偲屇偽傟偰偄傑偡丅 昦尨嬠偺PAMPs偲偟偰偼丄嵶嬠偺傋傫栄僞儞僷僋幙傗儁僾僠僪僌儕僇儞丄恀嬠偺僉僠儞偑傛偔尋媶偝傟偰偄傑偡丅
丂堦曽丄嵶朎幙偵懚嵼偡傞昦尨嬠擣幆庴梕懱偼丄妀巁寢崌晹埵(NB: Nucleotide Binding)偲LRR傪傕偪丄NB-LRR宆柶塽庴梕懱偲屇偽傟偰偄傑偡丅 摦暔偱偼丄NB僪儊僀儞偼Nod (Nucleotide oligomerization domain)偲屇偽傟偰偄傑偡偑丄嬤擭偱偼丄NLR (NB:NOD-like receptor)偲憤徧偝傟傞偙偲偑 懡偔側傝傑偟偨丅NB-LRR宆庴梕懱偼丄昦尨嬠偑怉暔偺柶塽墳摎傪慾奞偡傞偨傔偵暘斿偡傞僄僼僃僋僞乕偲憤徧偝傟傞僞儞僷僋幙傪専弌偟丄 柶塽墳摎傪桿摫偡傞偙偲偑抦傜傟偰偄傑偡乮僄僼僃僋僞乕桿摫惈柶塽乯丅恾侾偱偼丄堦晹偺嵶嬠偑僄僼僃僋僞乕傪暘斿偡傞偨傔偵巊梡偡傞 Type III暘斿僔僗僥儉傪帵偟偰偄傑偡偑丄 昦尨嬠乮嵶嬠丄恀嬠丄棏嬠側偳乯偺庬椶偵傛偭偰暘斿偡傞僔僗僥儉偼堎側偭偰偄傞偙偲偑抦傜傟偰偄傑偡丅
丂PRR偵傛傞PAMP擣幆偵敽偭偰丄峈嬠惈僞儞僷僋幙偺惗惉傗僇儘乕僗偵傛傞嵶朎暻偺嫮壔側偳條乆側杊屼斀墳偑桿摫偝傟傑偡丅 偙傟傜偺杊屼斀墳傪桿摫偡傞怣崋揱払偵偍偄偰丄MAP僉僫乕僛僇僗働乕僪乮MAPKKK丄MAPKK丄MAPK偺俁偮偺儕儞巁壔峺慺偐傜惉傞怣崋揱払宱楬乯 偑庡梫側栶妱傪壥偨偟偰偄傞偲峫偊傜傟偰偄傑偡丅偟偐偟丄MAPK僇僗働乕僪偺妶惈壔婡峔偍傛傃MAPK傪夘偟偨柶栶桿摫婡峔偼偁傑傝傢偐偭偰偄傑偣傫丅 堦曽丄NB-LRR宆庴梕懱偵傛偭偰桿摫偝傟傞僄僼僃僋僞乕桿摫惈柶塽偼丄懡偔偺応崌丄嵶朎巰傪敽偆嫮偄掞峈惈斀墳傪桿摫偟傑偡丅屆偔偐傜懴昦惈堢庬偺偨傔丄 昦婥偵懳偟偰嫮偄懴惈傪晅梌偡傞掞峈惈堚揱巕嵗偑堚揱妛揑偵摨掕偝傟丄堢庬慺嵽偺曣杮偲偟偰巊梡偝傟偰偒傑偟偨偑丄尰嵼偱偼丄 偦傟傜偺掞峈惈堚揱巕嵗偺懡偔偑NB-LRR宆庴梕懱傪僐乕僪偟偰偄傞偙偲偑柧傜偐偵側偭偰偄傑偡丅 偟偐偟丄NB-LRR宆庴梕懱偑偳偺傛偆偵柶塽斀墳傪桿摫偟偰偄傞偐偵偮偄偰偼偁傑傝棟夝偝傟偰偄傑偣傫丅
尋媶撪梕
1)怉暔偺僷僞乕儞桿摫惈柶塽偺暘巕婡峔偺夝柧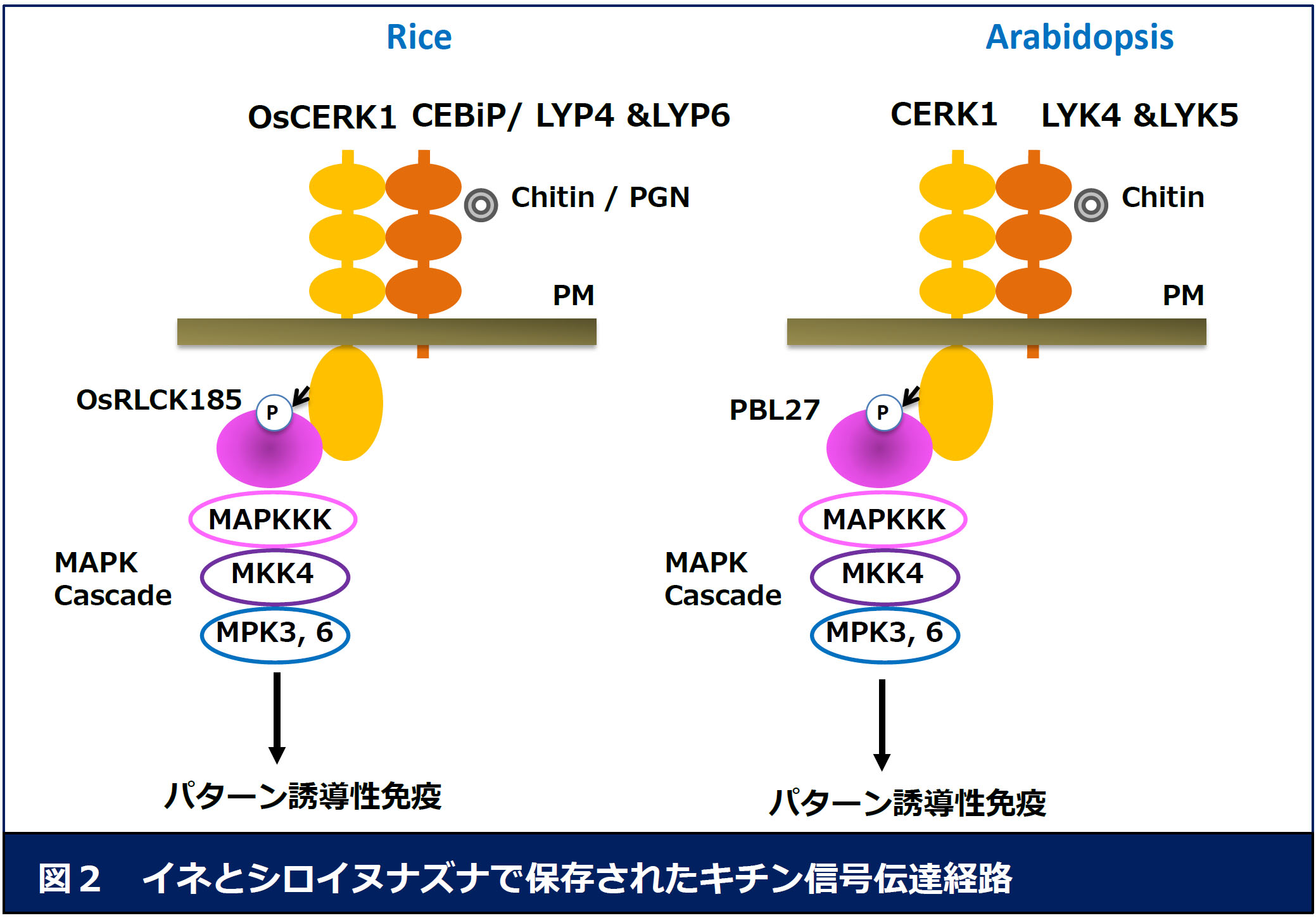 丂嵶朎枌忋偵懚嵼偡傞僷僞乕儞擣幆庴梕懱(PRR)偼丄PAMPs傪擣幆偟偰嵶朎撪偵杊屼斀墳傪桿摫偡傞偙偲偑抦傜傟偰偄傑偡偑丄偦偺徻嵶偼柧傜偐偵側偭偰偄傑偣傫丅
変乆偼丄恀嬠偺PAMP偱偁傞僉僠儞傗昦尨嵶嬠偺PAMP偱偁傞儁僾僠僪僌儕僇儞(PGN)偺擣幆偵傛偭偰桿摫偝傟傞柶塽宯傪儌僨儖偲偟偰尋媶傪恑傔偰偄傑偡丅偙傟傑偱偵丄
僉僠儞偲寢崌偡傞RLP偲偟偰CEBiP偑摨掕偝傟丄PGN傪擣幆偡傞RLP偲偟偰LYP4/LYP6偑曬崘偝傟偰偄傑偡丅偙傟傜偺RLP偑僉僠儞/PGN傪擣幆偡傞偲丄
RLK偱偁傞OsCERK1偲暋崌懱傪宍惉偟丄偦傟偑堷偒嬥偲側偭偰嵶朎撪偵杊屼斀墳偑桿摫偝傟傞偙偲偑抦傜傟偰偄傑偡丅変乆偼丄偙傟傑偱偵Receptor-like
cytoplasmic kinase (RLCK)僼傽儈儕乕偵懏偡傞OsRLCK185傪扨棧偟丄OsRLCK185偑OsCERK1偺嵶朎撪僉僫乕僛僪儊僀儞偲憡屳嶌梡偡傞偙偲丄偝傜偵僉僠儞擣幆偵
敽偭偰OsCERK1偑OsRLCK185傪儕儞巁壔偡傞偙偲傪柧傜偐偵偟傑偟偨乮恾俀乯丅傑偨丄OsRLCK185敪尰梷惂懱偱偼丄僉僠儞偍傛傃PGN偵傛偭偰桿摫偝傟傞杊屼斀墳偑梷惂偝傟傞偙偲偐傜丄OsRLCK185偑PGN偲僉僠儞偺擣幆偵娭傢傞OsCERK1偺怣崋揱払宯偱摥偄偰偄傞偙偲偑柧傜偐偵側傝傑偟偨丅杮惉壥偼丄Cell
Host & Microbe 2013擭3寧崋偱曬崘偟傑偟偨偑(Yamaguchi et al 2013)丄宖嵹崋偺Featured
Article偵慖偽傟丄崙撪偱傕懡偔偺儊僨傿傾偵庢傝忋偘偰傕傜偄傑偟偨丅
丂嵶朎枌忋偵懚嵼偡傞僷僞乕儞擣幆庴梕懱(PRR)偼丄PAMPs傪擣幆偟偰嵶朎撪偵杊屼斀墳傪桿摫偡傞偙偲偑抦傜傟偰偄傑偡偑丄偦偺徻嵶偼柧傜偐偵側偭偰偄傑偣傫丅
変乆偼丄恀嬠偺PAMP偱偁傞僉僠儞傗昦尨嵶嬠偺PAMP偱偁傞儁僾僠僪僌儕僇儞(PGN)偺擣幆偵傛偭偰桿摫偝傟傞柶塽宯傪儌僨儖偲偟偰尋媶傪恑傔偰偄傑偡丅偙傟傑偱偵丄
僉僠儞偲寢崌偡傞RLP偲偟偰CEBiP偑摨掕偝傟丄PGN傪擣幆偡傞RLP偲偟偰LYP4/LYP6偑曬崘偝傟偰偄傑偡丅偙傟傜偺RLP偑僉僠儞/PGN傪擣幆偡傞偲丄
RLK偱偁傞OsCERK1偲暋崌懱傪宍惉偟丄偦傟偑堷偒嬥偲側偭偰嵶朎撪偵杊屼斀墳偑桿摫偝傟傞偙偲偑抦傜傟偰偄傑偡丅変乆偼丄偙傟傑偱偵Receptor-like
cytoplasmic kinase (RLCK)僼傽儈儕乕偵懏偡傞OsRLCK185傪扨棧偟丄OsRLCK185偑OsCERK1偺嵶朎撪僉僫乕僛僪儊僀儞偲憡屳嶌梡偡傞偙偲丄偝傜偵僉僠儞擣幆偵
敽偭偰OsCERK1偑OsRLCK185傪儕儞巁壔偡傞偙偲傪柧傜偐偵偟傑偟偨乮恾俀乯丅傑偨丄OsRLCK185敪尰梷惂懱偱偼丄僉僠儞偍傛傃PGN偵傛偭偰桿摫偝傟傞杊屼斀墳偑梷惂偝傟傞偙偲偐傜丄OsRLCK185偑PGN偲僉僠儞偺擣幆偵娭傢傞OsCERK1偺怣崋揱払宯偱摥偄偰偄傞偙偲偑柧傜偐偵側傝傑偟偨丅杮惉壥偼丄Cell
Host & Microbe 2013擭3寧崋偱曬崘偟傑偟偨偑(Yamaguchi et al 2013)丄宖嵹崋偺Featured
Article偵慖偽傟丄崙撪偱傕懡偔偺儊僨傿傾偵庢傝忋偘偰傕傜偄傑偟偨丅 丂偝傜偵丄OsRLCK185偺僔儘僀僰僫僘僫儂儌儘僌偲偟偰PBL27傪扨棧偟傑偟偨丅 PBL27傕CERK1偲寢崌偟丄僉僠儞擣幆偵敽偭偰丄CERK1偵儕儞巁壔傪庴偗傞偙偲偑柧傜偐偲側傝丄扨巕梩偱偁傞僀僱偲憃巕梩偱偁傞僔儘僀僰僫僘僫偺娫偱曐懚偝傟偨 僉僠儞怣崋揱払宱楬偑懚嵼偡傞偙偲偑柧傜偐偵側傝傑偟偨乮恾俀乯丅杮尋媶偼丄柧帯戝丒廰扟尋偲偺嫟摨尋媶偱峴傢傟傑偟偨(Shinya and Yamaguchi et al. Plant J 2014)丅 尰嵼偼丄僉僠儞擣幆婡峔偲嵶朎撪柶塽僔僌僫儖偺妶惈壔婡峔偺慡杄夝柧偵岦偗丄PBL27傪拞怱偵丄CERK1暋崌懱偺 峔惉場巕偺婡擻夝愅傪恑傔偰偄傑偡丅
俀乯昦尨嬠僄僼僃僋僞乕傪棙梡偟偨怉暔柶塽場巕偺摨掕偲婡擻夝愅
 丂昦尨嬠偼丄帺恎偺僞儞僷僋幙乮埲壓丄僄僼僃僋僞乕乯傪怉暔偺嵶朎撪偵憲傝崬傒丄怉暔偺柶塽墳摎傪慾奞偟傑偡丅 僄僼僃僋僞乕傪怉暔嵶朎撪偱敪尰偝偣傞偲丄怉暔偺柶塽墳摎偑嫮偔慾奞偝傟傞偙偲偐傜丄僄僼僃僋僞乕偼柶塽桿摫偵廳梫側僞儞僷僋幙
偺摥偒傪慾奞偟偰偄傞偲峫偊傜傟傑偡丅偦偙偱丄変乆偼丄僀僱偺廳梫昦奞偱偁傞敀梩屚昦嬠偺僄僼僃僋僞乕傪棙梡偟偰丄 偦傟傜偑昗揑偲 偡傞怉暔柶塽場巕傪扵嶕偡傞偙偲偱丄怉暔柶塽偵偍偄偰廳梫側摥偒傪傕偮僞儞僷僋幙傪摨掕偟偰偄傑偡丅偝傜偵丄摨掕偟偨怉暔柶塽場巕
偺婡擻傪夝愅偡傞偙偲偱丄怉暔柶塽偺怣崋揱払宯傪柧傜偐偵偟偰偄傑偡丅
丂昦尨嬠偼丄帺恎偺僞儞僷僋幙乮埲壓丄僄僼僃僋僞乕乯傪怉暔偺嵶朎撪偵憲傝崬傒丄怉暔偺柶塽墳摎傪慾奞偟傑偡丅 僄僼僃僋僞乕傪怉暔嵶朎撪偱敪尰偝偣傞偲丄怉暔偺柶塽墳摎偑嫮偔慾奞偝傟傞偙偲偐傜丄僄僼僃僋僞乕偼柶塽桿摫偵廳梫側僞儞僷僋幙
偺摥偒傪慾奞偟偰偄傞偲峫偊傜傟傑偡丅偦偙偱丄変乆偼丄僀僱偺廳梫昦奞偱偁傞敀梩屚昦嬠偺僄僼僃僋僞乕傪棙梡偟偰丄 偦傟傜偑昗揑偲 偡傞怉暔柶塽場巕傪扵嶕偡傞偙偲偱丄怉暔柶塽偵偍偄偰廳梫側摥偒傪傕偮僞儞僷僋幙傪摨掕偟偰偄傑偡丅偝傜偵丄摨掕偟偨怉暔柶塽場巕
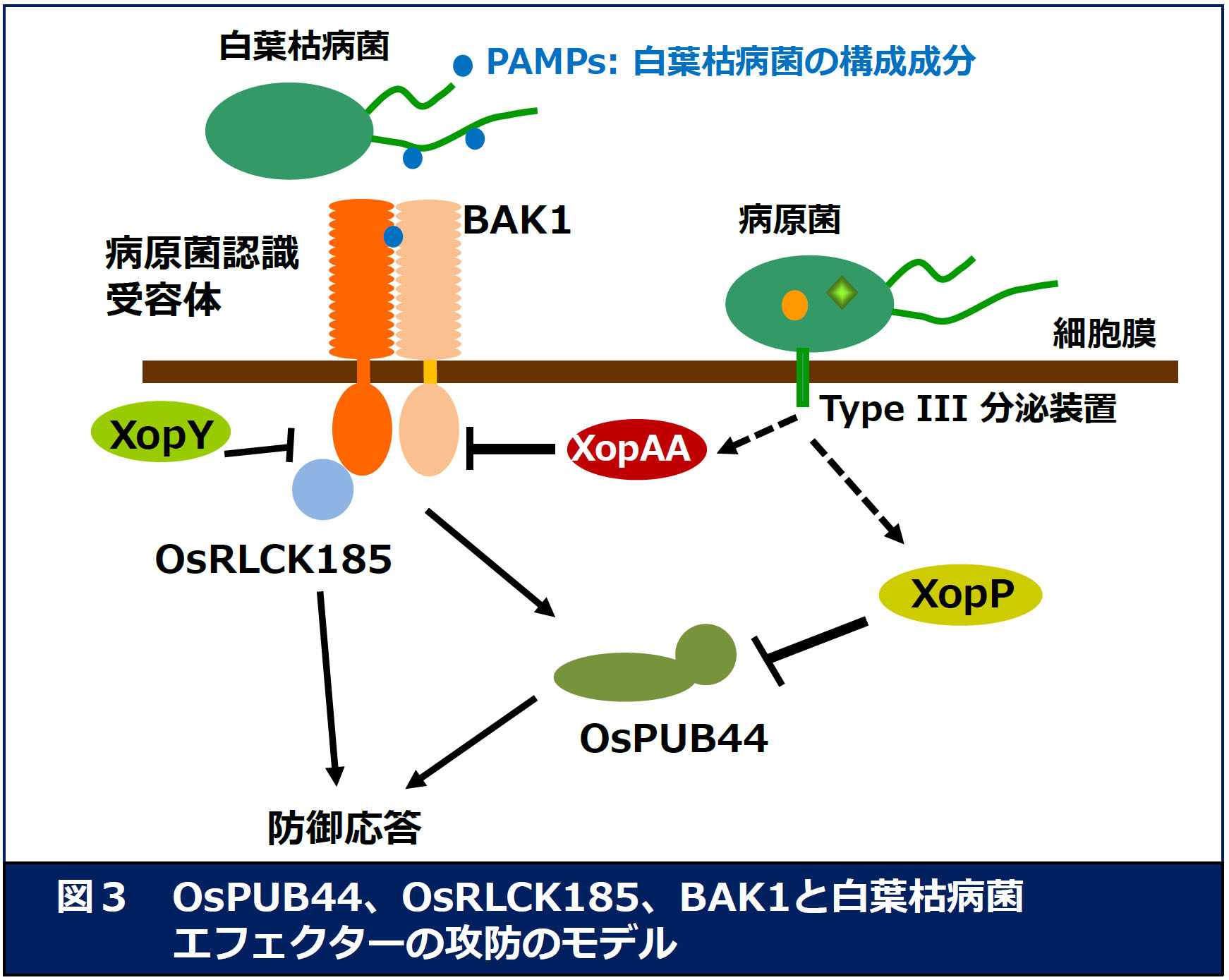
偺婡擻傪夝愅偡傞偙偲偱丄怉暔柶塽偺怣崋揱払宯傪柧傜偐偵偟偰偄傑偡丅 丂偙傟傑偱偵変乆偼丄僀僱偺敀梩屚昦嬠偺僄僼僃僋僞乕XopY (Xoo1488)偑昗揑偲偟偰偄傞僀僱偺場巕傪扵嶕偟丄OsRLCK185傪扨棧偟傑偟偨 乮恾俁乯丅夝愅偺寢壥丄XopY偼丄CERK1偵傛傞OsRLCK185偺儕儞巁壔傪慾奞偡傞偙偲偑柧傜偐偵側傝傑偟偨 (Yamaguchi et al. Cell Host Microbe 2013)丅偝傜偵丄僀僱偺敀梩屚昦嬠偺僄僼僃僋僞乕XopP(Xoo3222)傪夝愅偟 丄憡屳嶌梡場巕偲偟偰OsPUB44傪摨掕偟傑偟偨丅OsPUB44偼丄U-box僪儊僀儞傪傕偮儐價僉僠儞E3儕僈乕僛偱偁傝丄 柶塽墳摎偺億僕僥傿僽儗僊儏儗乕僞乕偲偟偰婡擻偟偰偄傞偙偲偑傢偐傝傑偟偨丅偝傜偵丄XopP偼OsPUB44偺U-box偵寢崌偡傞偙偲偱丄 儕僈乕僛妶惈傪慾奞偟丄柶塽墳摎傪梷惂偟偰偄傞偙偲偑柧傜偐偵側傝傑偟偨丅杮惉壥偼丄Nature Communications偵敪昞偟丄 崙撪偵偍偄偰傕懡偔偺儊僨傿傾偵庢傝忋偘傜傟傑偟偨丅
丂尰嵼丄XopY傗XopP傪娷傓敀梩屚昦嬠僄僼僃僋僞乕偺婡擻夝愅傪恑傔傞偲偲傕偵丄OsPUB44偺憡屳嶌梡場巕偺扨棧偵傛偭偰OsPUB44傪 夘偟偨柶栶桿摫宯偺夝愅傪峴偭偰偄傑偡丅
俁乯怉暔偺僄僼僃僋僞乕桿摫惈柶塽偺暘巕婡峔偺夝柧
丂丂NB-LRR宆庴梕懱偼丄僄僼僃僋僞乕傪擣幆偡傞偙偲偵傛傝丄嫮偄掞峈惈斀墳傪桿摫偟傑偡偑丄偦偺暘巕婡峔偼偁傑傝柧傜偐偵側偭偰偄傑偣傫丅 僀僱敀梩屚昦嬠偺掞峈惈堚揱巕Xa1偼丄NB-LRR宆庴梕懱偱偡偑 (Yoshimura et al. PNAS 1998)丄偳偺傛偆側僄僼僃僋僞乕傪擣幆偟丄偳偺傛偆偵柶塽墳摎傪桿摫 偟偰偄傞偐偵偮偄偰偼柧傜偐偵側偭偰偄傑偣傫丅偦偙偱丄Xa1傪惂屼偡傞敀梩屚昦嬠偺僄僼僃僋僞乕偍傛傃Xa1偲憡屳嶌梡偡傞僀僱柶塽場巕偺夝愅傪恑傔偰偍傝丄 Xa1傪夘偟偨柶栶桿摫婡峔偺慡杄傪柧傜偐偵偡傞偙偲傪栚巜偟偰尋媶偟偰偄傑偡丅
係)怉暔柶塽偵偍偗傞MAP僉僫乕僛僇僗働乕僪偺妶惈壔婡峔偺夝愅
怉暔柶塽偺弶婜墳摎偵偍偄偰丄MAP僉僫乕僛(MAPK)僇僗働乕僪偑妶惈壔偝傟傞偙偲偑傛偔抦傜傟偰偄傑偡丅偟偐偟丄庴梕懱偺壓棳偱丄 偳偺傛偆偵MAPK僇僗働乕僪偑妶惈壔偝傟傞偐偵偮偄偰偼慡偔柧傜偐偵側偭偰偄傑偣傫丅忋婰偺OsRLCK185傗PBL27偺夝愅偵傛傝丄OsRLCK185傗PBL27偑MAPK僇僗働乕僪 偺妶惈壔傪惂屼偟偰偄傞偙偲偑傢偐傝傑偟偨丅尰嵼丄OsRLCK185傗PBL27偑捈愙憡屳嶌梡偡傞MAPKKK傪摨掕偟丄偦偺夝愅傪捠偠偰丄RLCK僼傽儈儕乕偵傛傞MAPK 僇僗働乕僪偺妶惈壔婡峔傪夝柧偡傞偙偲傪栚巜偟偰偄傑偡丅
俆) 怉暔柶塽桿摫偁傞偄偼昦尨嬠偺昦尨惈傪梷惂偡傞暔幙偺扵嶕偲夝愅 丂
怉暔柶塽尋媶偵傛偭偰摼傜傟偨婎慴揑抦尒傪傕偲偵丄怴偟偄僞僀僾偺擾栻傪扵嶕偡傞偨傔偺僗僋儕乕僯儞僌宯傪峔抸偟 丄怉暔柶塽傪桿摫偡傞條乆側壔崌暔傗丄 昦尨嬠偺昦尨惈偺傒傪梷惂偡傞壔崌暔側偳丄娐嫬偵傗偝偟偄擾栻偺奐敪傪栚巜偟偨尋媶傪峴偭偰偄傑偡丅
俇) 娐嫬偵傗偝偟偄懴昦惈媄弍偺奐敪
丂柶塽斀墳偵偍偄偰丄峈嬠惈暔幙偺拁愊丄妶惈巁慺惗惉丄僇儘乕僗偺宍惉丄昦尨嬠傪暘夝偡傞峺慺偺敪尰側偳丄 條乆側杊屼僔僗僥儉偑妶惈壔偝傟傑偡偑丄偦傟傜偺巒揰偲側傞偺偼昦尨嬠擣幆偱偡丅変乆偼丄昦尨嬠擣幆婡峔偍傛傃偦傟偵 懕偔怣崋揱払婡峔偺婎慴尋媶偍傛傃偦偺棟夝傪捠偟偰丄偙偺傛偆側懡柺揑側杊屼斀墳傪嫤挷揑偵桿摫偱偒傞懴昦惈媄弍偺峔抸傪 栚巜偟偰偄傑偡丅嬤擭丄僔儘僀僰僫僘僫傗僀僱偱摼傜傟偨堚揱巕偁傞偄偼忣曬傪丄懠偺怉暔丒嶌暔偵墳梡偡傞 乽Translational Research乿偑幚尰偟偰偄傑偡偑丄 変乆偺尋媶惉壥偑墳梡媄弍偲偟偰丄怘椘惗嶻傗僄僱儖僊乕惗嶻偵峷專偱偒傞偙偲傪婜懸偟偰偄傑偡丅
仚仚仚
仚仚
嬤婨戝妛擾妛晹
僶僀僆僒僀僄儞僗妛壢
怉暔暘巕堚揱妛嫵幒
價儖僟乕僋儕僯僢僋
仹631-8505
撧椙導撧椙巗拞挰3327-204
TEL 0742-43-7335
FAX 0742-43-7335